Митохондрии краткое определение. Митохондрии – строение и функции
1 - наружная мембрана;
3 - матрикс;
2 - внутренняя мембрана;
4 - перимитохондриальное пространство.
Свойства митохондрий (белки, структура) закодированы частично в ДНК митохондрий, а частично в ядре. Так, митохондриальный геном кодирует белки рибосом и частично систему переносчиков электронотранспортной цепи, а в геноме ядра кодирована информация о белках-ферментах цикла Кребса. Сопоставление размеров митохондриальной ДНК с числом и размером мито-хондриальных белков показывает, что в ней заложено информации почти для половины белков. Это и позволяет считать митохондрии, как и хлоропласты, полуавтономными, т. е. не полностью зависящими от ядра. Они имеют собственную ДНК и собственную белоксинтезирующую систему, и именно с ними и с пластидами связана так называемая цитоплазматическая наследственность. В большинстве случаев это наследование по материнской линии, так как инициальные частицы митохондрий локализованы в яйцеклетке. Таким образом, митохондрии всегда образуются от митохондрий. Широко обсуждается вопрос, как рассматривать митохондрии и хлоропласты с эволюционной точки зрения. Еще в 1921 г. русский ботаник Б.М. Козо-Полянский высказал мнение, что клетка - это симбиотрофная система, в которой сожительствует несколько организмов. В настоящее время эндосимбиотическая теория происхождения митохондрий и хлоропластов является общепринятой. Согласно этой теории, митохондрии - это в прошлом самостоятельные организмы. По мнению Л. Маргелис (1983), это могли быть эубактерии, содержащие ряд дыхательных ферментов. На определенном этапе эволюции они внедрились в примитивную, содержащую ядро, клетку. Оказалось, что ДНК митохондрий и хлоропластов по своей структуре резко отличается от ядерной ДНК высших растений и сходна с бактериальной ДНК (кольцевое строение, нуклеотидная последовательность). Сходство обнаруживается и по величине рибосом. Они мельче цитоплазматических рибосом. Синтез белка в митохондриях, подобно бактериальному, подавляется антибиотиком хлорамфениколом, который не влияет на синтез белка на рибосомах эукариот. Кроме того, система переноса электронов у бактерий расположена в плазматической мембране, что напоминает организацию электронтранспортной цепи во внутренней митохондриальной мембране.
Митохондрия (с греческого μίτος (митос) – нить и χονδρίον (хондрион) – гранула) клеточная – двумембранный органоид, содержит свой собственный генетический материал, митохондриальную . Они встречаются как сферические или трубчатые клеточные структуры у почти всех эукариотов, но не у прокариотов.
Митохондрии – это органеллы, которые регенерируют высокоэнергетическую молекулу аденозинтрифосфата через дыхательную цепь. В дополнение к этому окислительному фосфорилированию они выполняют другие важные задачи, например, участвуют в образовании кластеров железа и серы . Строение и функции таких органоидов подробно рассмотрены ниже.
Вконтакте
Общие сведения
Особенно много находится митохондрий в с высоким энергопотреблением. К ним относятся мышечные, нервные, сенсорные клетки и ооциты. В клеточных структурах сердечной мышцы объемная доля этих органоидов достигает 36 %. Они имеют диаметр около 0.5-1.5 мкм и разнообразные формы, от сфер до сложных нитей. Их число корректируется с учетом энергетических потребностей клетки.
Эукариотические клетки, которые теряют свои митохондрии, не могут их восстановить . Существуют также эукариоты без них, например, некоторые простейшие. Количество данных органоидов на клеточную единицу обычно составляет от 1000 до 2000 при объемной доле в 25 %. Но эти значения могут сильно варьироваться в зависимости от типа клеточной структуры и организма. В зрелой клетке спермы их около четырех-пяти, в зрелой яйцеклетке – несколько сотен тысяч.
Митохондрии передаются через плазму яйцеклетки только от матери, что стало причиной исследования материнских линий. В настоящее время установлено, что также через сперму некоторые мужские органоиды импортируются в плазму оплодотворенной яйцеклетки (зиготы). Вероятно, они будут устранены довольно быстро. Однако есть несколько случаев, когда врачи смогли доказать, что митохондрии ребенка были отцовской линии. Заболевания, вызванные мутациями в митохондриальных генах, наследуются только от матери.
Интересно! Популярный научный термин «энергетическая станция клетки» был придуман в 1957 году Филиппом Сикевицем.
Схема строения митохондрии
Рассмотрим особенности строения этих важных структур. Они образованы в результате сочетания нескольких элементов. Оболочка этих органоидов складывается из внешней и внутренней мембраны, они в свою очередь состоят из фосфолипидных бислоев и белков. Обе оболочки отличаются по своим свойствам. Между ними расположено пять различных отсеков: наружная мембрана, межмембранное пространство (промежуток между двумя мембранами), внутренняя, криста и матрикс (пространство внутри внутренней мембраны), в целом – внутренние структуры органоида.
На иллюстрациях в учебниках митохондрия преимущественно выглядит как отдельная бобовидная органелла. Так ли это на самом деле? Нет, они образуют трубчатую митохондриальную сеть , которая может проходить и изменять всю клеточную единицу. Митохондрии в клетке способны сочетаться (путем слияния) и повторно делиться (делением).
Обратите внимание! В дрожжах за одну минуту совершается около двух митохондриальных слияний. Поэтому невозможно точное определение текущей численности митохондрий в клетках.
Внешняя мембрана
 Наружная оболочка окружает всю органеллу и включает в себя каналы белковых комплексов, что позволяют обмен молекулами и ионами между митохондрией и цитозолем. Крупные молекулы не могут пройти через мембрану
.
Наружная оболочка окружает всю органеллу и включает в себя каналы белковых комплексов, что позволяют обмен молекулами и ионами между митохондрией и цитозолем. Крупные молекулы не могут пройти через мембрану
.
Внешняя, которая охватывает всю органеллу и не свернута, имеет весовое отношение фосфолипида к белку 1:1 и, таким образом, похожа на эукариотическую плазматическую мембрану. Она содержит множество интегральных белков, поринов. Порины образуют каналы, которые обеспечивают свободную диффузию молекул с массой до 5000 дальтон через оболочку. Более крупные белки могут вторгаться, когда сигнальная последовательность на N-конце связывается с большой субъединицей белка транслоксазы, из которой они затем активно перемещаются по мембранной оболочке.
Если трещины возникают во внешней оболочке, белки из межмембранного пространства могут выходить в цитозоль, что может привести к гибели клетки . Наружная мембрана может сливаться с оболочкой эндоплазматического ретикулума, а затем формировать структуру под названием MAM (ER, ассоциированную с митохондрией). Это важно для обмена сигналами между ER и митохондрией, что также необходимо для переноса .
Межмембранное пространство
Участок представляет собой промежуток посреди внешней и внутренней мембраны. Поскольку внешняя обеспечивает свободное проникновение малых молекул, их концентрация, таких как ионы и сахар, в межмембранном пространстве идентична концентрациям в цитозоле. Однако для больших белков требуется передача специфической сигнальной последовательности, так что состав белков различается между межмембранным пространством и цитозолем. Таким образом, белок, который удерживается в межмембранном промежутке, является цитохромом.
Внутренняя мембрана
Внутренняя митохондриальная мембрана содержит белки с четырьмя видами функций:
- Белки – проводят реакции оксидации респираторной цепочки.
- Аденозинтрифосфатсинтаза, которая производит в матрице АТФ.
- Специфические транспортные белки, которые регулируют проход метаболитов между матрицей и цитоплазмой.
- Системы импорта белков.
Внутренняя имеет, в частности, двойной фосфолипид, кардиолипин, замещенный четырьмя жирными кислотами. Кардиолипин обычно характерен для митохондриальных мембран и бактериальных плазматических мембран. В организме человека он в основном присутствует в областях с высокой метаболической активностью или высокой энергетической активностью, таких как сократительные кардиомиоциты, в миокарде.
Внимание! Внутренняя мембрана содержит более 150 различных полипептидов, около 1/8 всех митохондриальных белков. В результате концентрация липидов ниже, чем у внешнего бислоя, и его проницаемость ниже.
 Разделяется на многочисленные кристы, они расширяют внешнюю область внутренней митохондриальной оболочки, поднимая ее способность вырабатывать АТФ.
Разделяется на многочисленные кристы, они расширяют внешнюю область внутренней митохондриальной оболочки, поднимая ее способность вырабатывать АТФ.
В типичной митохондрии печени, например, внешняя область, в частности кристы, примерно в пять раз превышает площадь наружной мембраны. Энергетические станции клеток, которые имеют более высокие потребности в АТФ, например, мышечные клетки, содержат больше крист, чем типичная митохондрия печени.
Внутренняя оболочка охватывает матрикс, внутреннюю жидкость митохондрии. Он соответствует цитозолю бактерий и содержит митохондриальную ДНК, ферменты цитратного цикла и их собственные митохондриальные рибосомы, которые отличаются от рибосом в цитозоле (но также и от бактерий). Межмембранное пространство содержит ферменты, которые могут фосфорилировать нуклеотиды под потреблением АТФ.
Функции
- Важные пути деградации: цитратный цикл, для которого пируват вводится из цитозоля в матрикс. Затем пируват декарбоксилируют пируватдегидрогеназой до ацетилкофермента А. Другим источником ацетилкофермента А является деградация жирных кислот (β-окисление), которая происходит в клетках животных в митохондриях, но в растительных – только в глиоксисомах и пероксисомах. С этой целью ацилкофермент А переносят из цитозоля путем связывания с карнитином через внутреннюю митохондриальную мембрану и превращают в ацетилкофермента А. Из него большинство восстановительных эквивалентов в цикле Кребса (также известный как цикл Кребса или цикл трикарбоновой кислоты), которые затем превращаются в АТФ в окислительной цепи.
- Окислительная цепь. Установлен электрохимический градиент между межмембранным пространством и митохондриальным матриксом, который служит для получения АТФ с помощью АТФ-синтазы, с помощью процессов переноса электронов и накопления протонов. Электроны и протоны, необходимые для создания градиента, получают путем окислительной деградации из питательных веществ (например, глюкозы), поглощаемых организмом. Первоначально гликолиз происходит в цитоплазме.
- Апоптоз (запрограммированная гибель клеток)
- Хранение кальция: благодаря способности абсорбировать ионы кальция и затем высвобождать их, митохондрии вмешиваются в гомеостаз клетки.
- Синтез кластеров железа-серы, требуемый, среди прочего, многими ферментами дыхательной цепи. Эта функция теперь считается существенной функцией митохондрий, т.е. как это причина, по которой почти все клетки полагаются на энергетические станции для выживания.
![]()
Матрикс
Это пространство, включенное во внутреннюю митохондриальную мембрану. Содержит около двух третей общего белка. Играет решающую роль в производстве АТФ с помощью синтазы АТФ, включенной во внутреннюю мембрану. Содержит высококонцентрированную смесь сотен различных ферментов (главным образом, участвующих в деградации жирных кислот и пирувата), митохондриально-специфических рибосом, передаточной РНК и нескольких копий ДНК митохондриального генома.
Данные органоиды имеют свой собственный геном, а также ферментативное оборудование, необходимое для осуществления собственного биосинтеза белка .
Митохондрия Что такое Митохондрия и её функции
Строение и функционирование митохондрий
Вывод
Таким образом, митохондриями называются клеточные электростанции, которые производят энергию и занимают ведущее место в жизни и выживаемости отдельной клетки в частности и живого организма в целом. Митохондрии – это неотъемлемая часть живой клетки, в том числе растительной, которые до конца еще не изучены. Особенно много митохондрий в тех клетках, которым требуется больше энергии.
Митохондрии (МТ) – одно из самых интересных мне направлений исследований. Объединение митохондрий с другой клеткой в ходе эндосимбиоза около 1,6 млрд лет назад стало основной всех многоклеточных эукариотов со сложной структурой. Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Лучшее обзорное исследование последнего времени по митохондриях – работа Вернера Кулбрандта «Структура и функция митохондриальных белковых комплексов мембраны ». Если вы знаете английский язык и интересуетесь устройством этих органелл, то настоятельно рекомендую к прочтению. Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Митохондрия кодирует сама только 13 белков, не смотря на наличие отдельной от клетки ДНК (мтДНК) и всего «производственного» цикла по транскрипции белков. Изолированная митохондрия какое-то время может сохранять композицию и функционировать.
Рисунок 1. Компоненты мембраны митохондриона. Внешняя мембрана отделяет митохондрию от цитоплазмы. Она окружает внутреннюю мембрану, которая отделяет межмембранное пространство от богатого белками центрального матрикса. Внутреннюю мембрану разделяют на внутреннюю пограничную мембрану и кристы. Две эти части непрерывны в местах крепления крист (cristae junction ). Кристы простираются более или менее глубоко в матрикс и являются основным место митохондриального преобразования энергии. Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Внешняя мембрана пористая и позволяет веществам из цитоплазмы проходить через нее. Внутренняя мембрана плотная, для ее пересечения нужны транспортные белки [Гилберт Линг обоснованно не согласен], непрерывность барьера позволяет иметь внутренней мембране электрохимический потенциал в -180 mV. У матрикса довольно большой pH (7,9-8). Еще раз углублюсь в Линга. Щелочной (выше 7) pH способствует более развернутой конформации белков . Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
мтДНК находится в нуклеотидах, которых примерно 1000 на клетку. Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Внутренняя мембрана образует инвагинации, называемые кристами, которые глубоко проникают в матрикс. Кристы определяют третий «отсек» митохондрий – просвет крист (cristae lumen). Мембраны кристы содержат большинство, если не все, полностью «собранные» комплексы цепи переноса электронов и АТФ-синтазы. Просвет кристы содержит большое количество маленького растворимого белкового переносчика электронов (цитохром с). Митохондриальные кристы, таким образом, основное место биологической конверсии энергии во всех не фотосинтетических эукариотах.
С кристами тоже много всего интересного. Оптические свойства кристы влияют на распространение и генерацию света в тканях. Я даже встречал идеи о том, что поверхность крист подобна (предположение) поверхностям топологических изоляторов (подразумевалась суперпроводимость без диссипации заряда).
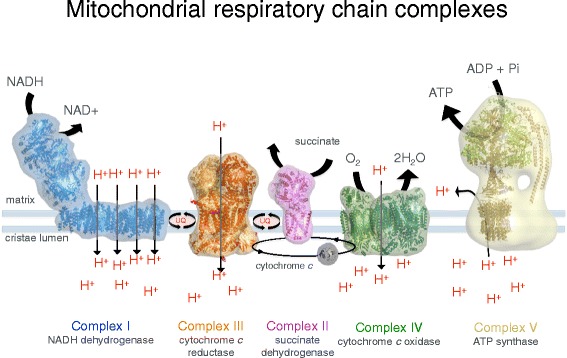
Рисунок 2. Мембранные белковые комплексы дыхательной цепи. Комплекс I (NADH / убихинон оксидоредуктаза, синий), Комплекс II (сукцинат дегидрогеназа, розовы), Комплекс III (цитохром С редуктаза, оранжевый), Комплекс IV (цитохром С оксидаза, зеленый) и митохондриальная АТФ синтаза (известная как комплекс V , бежевая) работают вместе во время окислительного фосфорилирования, чтобы клетки могли использовать энергию. Комплексы I , III , IV выкачивают протоны вдоль мембраны кристы, создавая протоновый градиент, стимулирующий синтез АТФ.
Теперь немного внимания на комплекс II. Вы помните, что жир (кето) делает упор в метаболизме на FADH2 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Еще прошу вас заметить, что комплекс II не выкачивает протоны. Что у нас рассеивает протоновый градиент, нарушает фосфорилирование и стимулирует сжигание жиров на тепло? Правильно, стресс холода. Термогенез связан с метаболизмом через комплекс, который не выкачивает протоны, тем самым не давая дополнительных протонов для АТФ-синтазы. Можно только удивляться как замечательно у нас продуман организм.
Крепления крист и MICOS
Места крепления крист (cristae junctions) – маленькие круглы отверстия примерно 25 нм диаметром. В митохондриях всех организмов есть система MICOS (mitochondria contact site and cristae to outer membrane), сборка из пяти мембранных и одного растворимого белков, прикрепляющих кристы к наружной мембране.
В клетках с повышенной потребностью в энергии, такие как скелетные и сердечные мышцы, кристы плотно заполняют большую часть объема митохондрии. В тканях с меньшими потребностями в энергии, таких как печень и почки, кристы находятся не так плотно по отношению друг к другу. Остается больше места в матрице для биосинтетических ферментов.
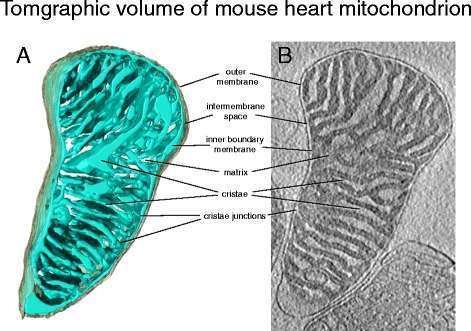
Рисунок 3. Томографический объем митохондрии сердца мыши. А) Трехмерный объем митохондрии сердца мыши, снятый cryo-ET. Наружная мембрана (серая) окутывает внутреннюю мембрану (светло-синяя). Внутренняя мембрана плотно наполнена кристами б) Томографический срез объема. Плотно заполненный матрикс, содержащий большую часть митохондриальных белков, выглядит темным на электронном микроскопе. В то время как межмембранное пространство и просветы крист выглядят светлыми из-за низкой концентрации белков.
Димеры АТФ синтазы
Митохондриальная F1-F0 АТФ синтаза является самым заметным белковым комплексом кристы. АТФ синтаза – это древняя наномашина, которая использует электрохимический протновых градиент вокруг внутренней мембраны для создания АФТ посредством вращательного катализа . Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Многие годы считалось, что АТФ синтаза случайным образом располагается на внутренней мембране. Но оказалось, что АТФ синтаза располагается двойными рядами . Причем линейные ряды АТФ синтазы – фундаментальный атрибут всех живых митохондрий.

Рисунок 4. Двойные ряды АТФ синтазы у семи разных видов.
Ряды АТФ синтазы располагаются в основном вдоль хребтов крист. Димеры изгибают липидный бислой и как следствие само-организуются в ряды. Когда у митохондрий дрожжей выбивали узлы e и g АФТ синтазы, то штамм рос на 60% медленней диких собратьев, и потенциал мембран их митохондрий был снижен вдвое. У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.

Рисунок 5. Структура димера АТФ синтазы митохондрии polymella sp. Вид сбоку на V-образный димер АТФ синтазы.
Комплексы и суперкомплексы дыхательной цепи
Протоновый градиент вокруг внутренней мембраны создается тремя крупными мембранными комплексами, известными как комплекс I, комплекс III и комплекс IV (см. рисунок 2). Комплекс I кормится электронами из NADH, высвобождаемая при передаче электрона энергия выкачивает четыре протона. Комплекс III получает электрон от восстановленного хинола и передает его носителю электронов (цитохрому с), выкачивая в процессе один протон. Комплекс IV получает электрон из цитохрома с и передает его молекулярному кислороду, выкачивая 4 протона за каждую молекулу кислорода, превращенную в воду. Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.

Рисунок 6. Комплекс I митохондрии коровьего сердца. Матриксная часть содержит ряд из восьми железно-серных (Fe-S) кластеров, которые направляют электроны из NADH в хинол на пересечении матрикса и мембраны. Мембранная часть состоит из 78 лопастей, включая выкачивающие протоны молекулы.
Комплексы I, III и IV соединяются в суперкомплексы или респирасомы. У пекарских дрожжей (saccharomyces cerevisiae ) нет комплекса I, их суперкомплексы состоят из III и IV. Роль суперкомплексов пока еще не ясна. Предполагают, что это делает транспорт электронов более эффективным, но прямых доказательств этому пока нет.

Рисунок 7. Суперкомплекс митохондрии коровьего сердца. Обратите внимание на дистанцию между комплексами I и III, который надо проделать хинолу. Стрелки – движения электрона в суперкомплексе.
Основным белком просвета кристы является цитохром с, который переносит электрон из комплекса III в комплекс IV. Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз .

Рисунок 8. Ряды димеров АТФ синтазы задают форму кристам. У хребта кристы АФТ синтаза (желтый) образует слив для протонов (красный), протоновые насосы электронной цепи (зеленый) находятся по обоим сторона рядов димеров. Направляя протоны от источника к АТФ синтазе, кристы работают как протоновые направляющие, позволяющие эффективное производство АТФ. Красные стрелки показывают направление потока протонов.
Реорганизация мембраны во время старения
Старение – фундаментальный и плохо понимаемый процесс всех эукариотов. Исследовали старение митохондрий на грибах Podospora anserina , которые живут всего 18 дней. В нормальной митохондрии кристы проникают глубоко в матрикс. Для этого нужны ряды димеров АТФ синтазы и MICOS комплекс у мест крепления крист. С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.
Транспорт электронов создает супероксид в комплексах I и III. Это побочный продукт метаболизма. Одновременно необходимый и смертельно опасный. Во время старения деление (fission) начинает превалировать над сращением (fussion). Это не дает поврежденным митохондриям «спастись» путем сращения и ускоряет неизбежное.

Рисунок 9. Изменения морфологии внутренней мембраны и димеров АТФ синтазы во время старения митохондрии.
Как видите, полей для будущих исследований очень много. Я предполагаю стык физики и биологии, где физики будут пытаются объяснить почему такая структура более энергетически эффективна. Тем более публикации по квантовой биологии






